Analytical Benchmark Solutions for the Cardiac EMI Model.
M.Sc. in Mathematics — University of Saskatchewan · August 2025
Advisor: Raymond J. Spiteri
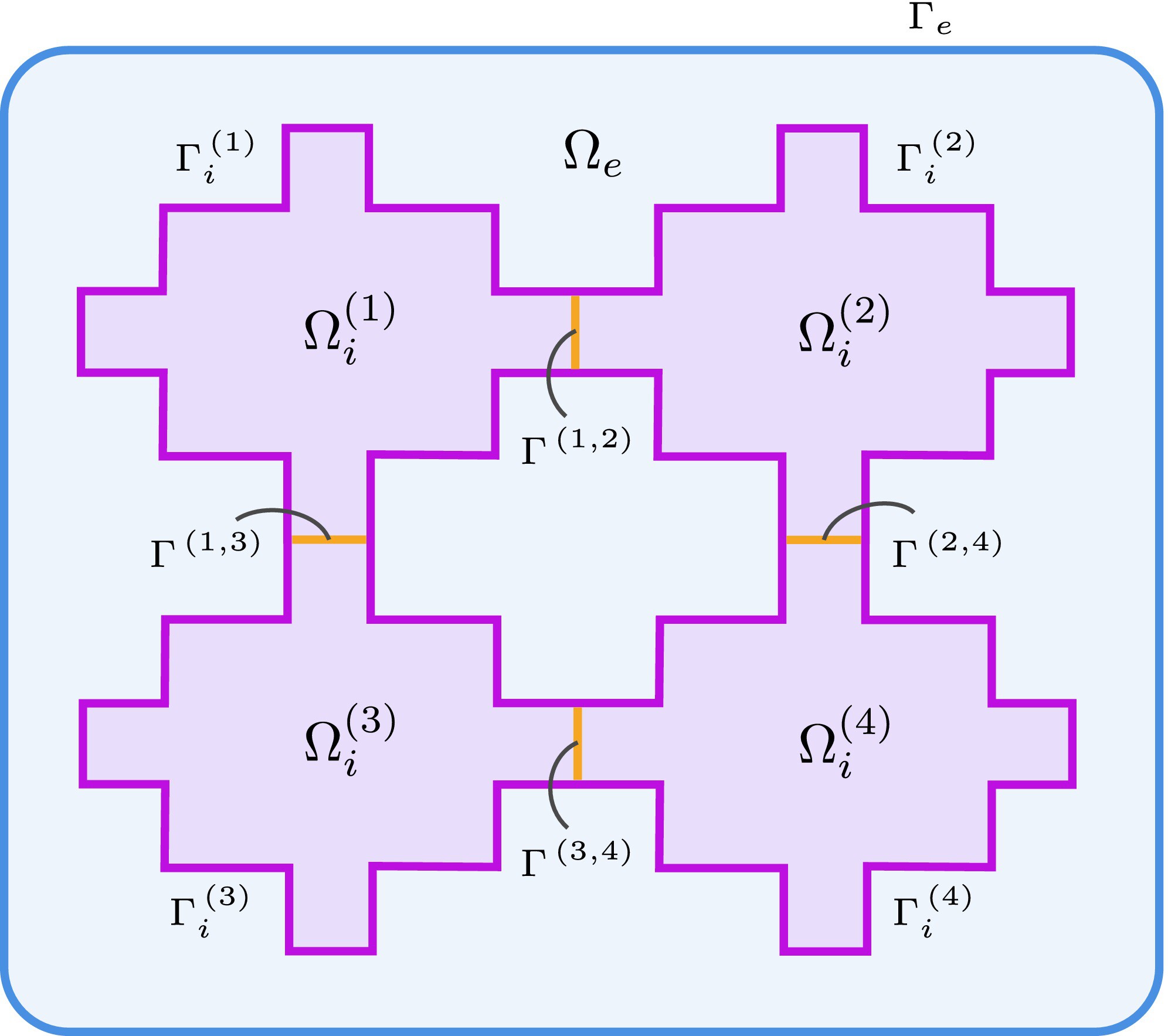
Figure 1. Configuration of a two-dimensional domain with four cardiac cells (\(\Omega_i^{(k)}\), purple) surrounded by extracellular space (\(\Omega_e\), blue) with gap junctions (\(\Gamma^{(k,\ell)}\), orange).
Abstract
This thesis develops analytical and manufactured benchmark solutions for the cardiac extracellular–membrane–intracellular (EMI) model, a cell-by-cell mathematical framework that explicitly resolves intracellular \((\Omega_i^{(k)})\), membrane \((\Gamma_i^{(k)})\), and extracellular \((\Omega_e)\) domains. Compared to traditional monodomain and bidomain models, the EMI model enables high-fidelity representation of cardiac dynamics, particularly under pathological conditions. Analytical solutions are derived for a single 2D circular cell in polar coordinates and for two coupled 3D semi-spherical cells in spherical coordinates, both under radial symmetry and equal intra- and extracellular conductivities. A manufactured solution for \(N\) cells in Cartesian coordinates enables comprehensive numerical validation via the mortar finite element method (MFEM) combined with operator-splitting (OS). A temporal convergence study confirms first-order accuracy for the Lie–Trotter OS method and second-order accuracy for the Strang OS method. The framework is applied to simulate cardiac channelopathies (Long QT1 and Brugada syndromes) using the ten Tusscher–Panfilov cell model.
The Cardiac EMI Model
The intracellular and extracellular potentials satisfy the following elliptic PDEs in their respective domains:
\[ \nabla \cdot \!\left(\sigma_i \nabla u_i^{(k)}\right) = 0 \quad \text{in } \Omega_i^{(k)}, \qquad \nabla \cdot \!\left(\sigma_e \nabla u_e\right) = 0 \quad \text{in } \Omega_e, \]where \(\sigma_i\) and \(\sigma_e\) are the intracellular and extracellular conductivities. The transmembrane potentials are defined as \(v^{(k)} = u_i^{(k)} - u_e\) on each membrane \(\Gamma_i^{(k)}\), and \(w^{(k,\ell)} = u_i^{(k)} - u_i^{(\ell)}\) across each gap junction \(\Gamma^{(k,\ell)}\). Their dynamics are governed by:
\[ \begin{aligned} C_m^{(k)} \frac{\partial v^{(k)}}{\partial t} &= I_m^{(k)} - I_{\mathrm{ion}}^{(k)}, &\quad& \text{on } \Gamma_i^{(k)}, \\[6pt] I_m^{(k)} &= \left(\sigma_e \nabla u_e\right)\cdot\hat{\mathbf{n}}_e = -\left(\sigma_i \nabla u_i^{(k)}\right)\cdot\hat{\mathbf{n}}_i^{(k)}, &\quad& \text{on } \Gamma_i^{(k)}, \\[6pt] C_m^{(k,\ell)} \frac{\partial w^{(k,\ell)}}{\partial t} &= I_m^{(k,\ell)} - I_{\mathrm{gap}}^{(k,\ell)}, &\quad& \text{on } \Gamma^{(k,\ell)}, \\[6pt] I_m^{(k,\ell)} &= \left(\sigma_i \nabla u_i^{(k)}\right)\cdot\hat{\mathbf{n}}_i^{(k)} = -\left(\sigma_i \nabla u_i^{(\ell)}\right)\cdot\hat{\mathbf{n}}_i^{(\ell)}, &\quad& \text{on } \Gamma^{(k,\ell)}, \end{aligned} \]where \(C_m^{(k)}\) is the membrane capacitance per unit area, \(I_{\mathrm{ion}}^{(k)}\) is the ionic current (given by a passive or active cell model), and \(\hat{\mathbf{n}}_i^{(k)}\) is the outward unit normal to \(\partial\Omega_i^{(k)}\). The extracellular potential \(u_e\) satisfies either a Dirichlet condition \(u_e = u_{\mathrm{app}}\) or a Neumann condition \((\sigma_e \nabla u_e)\cdot\hat{\mathbf{n}}_e = I_{\mathrm{app}}\) on \(\Gamma_e\).

Figure 2. Published paper in Mathematical Methods in the Applied Sciences, 2025.